1992 - A new Genus
MEXIPEDIUM: A NEW GENUS OF SLIPPER ORCHID
(CYPRIPEDIOIDEAE: ORCHIDACEAE)
Albert, V. & M.W. Chase
Lindleyana 7: 172-176, 1992
ABSTRACT
Mexipedium, based on Phragmipedium xerophyticum Soto, Salazar & Hagsater, is described as a new genus of slipper orchid (Cypripedioideae: Orchidaceae). It is segregated from Phragmipedium by its possession of unilocular rather than trilocular ovaries and can be distinguished from Paphiopedilum by its possession of branching racemes and valvate sepal aestivation. Phylogenetic analysis of anatomical, morphological, and molecular data indicates that Mexipedium, endemic to Oaxaca, Mexico, represents a basal branch of the New World, conduplicate-leaved lineage previously ascribed only to Phragmipedium.
THE RECENT description of Phragmipedium xerophyticum Soto, Salazar & Hagsater (Soto, Salazar, and Hagsater, 1990), a phenotypically and geographically isolated taxon from Oaxaca, Mexico, raises new problems in the taxonomic distinction between New World Phragmipedium Rolfe and Old World Paphiopedilum Pfitzer (Cypripedioideae: Orchidaceae). Paphiopedilum was erected to include all conduplicate-leaved slipper orchids (Pfitzer, 1886), only to have Phragmipedium segregated from it a decade later (Rolfe, 1896). Although distinction has been maintained through two nomenclatural conservations (Paphiopedilum-1959, Taxon 8: 242; Phragmipedium-1978, Taxon 27: 288) that have received global acceptance (CITES Plant Committee, 7th Meeting of the Conference Parties, Lausanne, Switzerland, October, 1989), the genera are similar anatomically and morphologically (Rosso, 1966; Atwood, 1984) and show few consistent differences. Phragmipedium xerophyticum presents a combination of Paphiopedilum-specific and Paphiopedilum-specific features. Its inclusion within Phragmipedium (Soto, Salazar, and Hagsater, 1990) was based upon the expression of four character states: (i) valvate vernation (aestivation) of the sepals, (ii) the absence of sinuous epidermal cells in the perianth, (iii) fusion of the lateral lobes of the labellum, and (iv) ventral synsepals larger than dorsal sepals (Atwood, 1984, p. 190). However, Phragmipedium xerophyticum and Paphiopedilum have unilocular ovaries, whereas all other Phragmipedium species have trilocular ovaries. The merit of these characters is discussed below with reference to the taxonomic status of Phragmipedium xerophyticum.
Sepal aestivation is not readily addressed for the entire Cypripedioideae. Rolfe (1896) recognized only valvate and imbricate aestivation, the latter confined to Paphiopedilum. Pfitzer (1903, p. 27) indicated that the calyx in Selenipedium Reichb.f. is characterized by "three sepals, free medially, connate laterally, aestivation lightly imbricate or perforate." In the most recent treatment of the subfamily Cypripedioideae, Atwood (1984) reported valvate aestivation for Phragmipedium, imbricate aestivation for Paphiopedilum, and perforate aestivation for both Cypripedium L. and Selenipedium. Despite confusion surrounding aestivation in Cypripedium and Selenipedium, it is clear that Phragmipedium may be distinguished consistently from Paphiopedilum by valvate versus imbricate aestivation.
Atwood reported (1984; without supporting figures or detailed discussion) the presence of sinuous-walled epidermal cells on the perianth parts of Cypripedium , Paphiopedilum , and Selenipedium, and their absence in Phragmipedium. In Cypripedium and Selenipedium, Atwood described these cells as similar to those of leaf epidermis. In Paphiopedilum, these sinuous-walled cells were observed only on or near the edges of perianth parts ( except in Paphiopedilum venustum [Wallich] Pfitzer ex Stein, where none were observed). Although "on or near" perianth margins is not a precise definition, we interpret the states of this character to be (i) absent, (ii) present marginally, or (iii) present medially and marginally. Nevertheless, the consistency of this trait is left in question by Atwood's (1984, p. 176) unqualified criterion "At least some epidermal cell walls sinuous" (our emphasis) and its polymorphism within Paphiopedilum.
Fusion of the lateral labellum lobes may be polymorphic within Phragmipedium. One of us (VAA) has observed this phenomenon extensively; however, specimens of Phragmipedium schlimii (Linden & Reichb.f.) Rolfe (voucher: L. Hegedus ex V. Albert VA89PH1, NCU) and Phragmipedium besseae Dodson & Kuhn (voucher: R. Topper ex V. Albert VA89PH7, NCU) examined have shown no signs of fusion. Indeed, Atwood (1984, p. 148) indicated a "nearly borderline character state" in his determination of labellum fusion for Phragmipedium schlimii. Although Soto, Salazar, and Hagsater (1990) stated that Phragmipedium xerophyticum has fused lateral lobes, this condition is not readily apparent in any of the accompanying photographs or drawings (see Fig. 1). G. Salazar (pers. comm.) found close proximity but not fusion of the lateral lobes in liquid preserved specimens at AMO. Phragmipedium xerophyticum, P. schlimii, and P. besseae all have simple, calceolate labella similar to those of Cypripedium, Selenipedium, and some Paphiopedilum species (e.g., Paphiopedilum delenatii Guillaumin); if fusion of the lateral lobes does occur in these taxa, it may be broken during or shortly after anthesis. Nevertheless, these observations indicate that this characteristic cannot be considered a uniform state marking Phragmipedium until careful ontogenetic studies are performed.
The last diagnostic feature for Phragmipedium, ventral synsepals larger than dorsal sepals, is a quantitative character that is variable across the Cypripedioideae, precluding precise designations of character states. Phragmipedium may, nevertheless, express a tendency in this particular direction (see Atwood, 1984, p. 145 , for area-ratio measurements of synsepal to dorsal sepal for Paphiopedilum and Phragmipedium species only).
The above critique of features considered diagnostic for Phragmipedium indicates that strong support for the naturalness (monophyly) of the genus has not been shown. Relative to Paphiopedilum, only the first Phragmipedium-specific characteristic discussed (valvate sepal aestivation) permits a clear distinction between the genera; the absence of sinuous-walled epidermal cells, labellum fusion, and relative sepal size do not seem as consistent. A more robust charactenstic, although not uniform within Phragmipedium, is the presence of branched racemes in Phragmipedium xerophyticum and several other species, which contrasts with the uniformly unbranched racemes of Paphiopedilum (Atwood, 1984; Soto, Salazar, and Hagsater, 1990; V. A. Albert, pers. obsv.).
Ovary placentation has been used as a fundamental characteristic for generic recognition within the Cypripedioideae (Reichenbach, 1854; Pfitzer, 1903; Rolfe, 1896). Phragmipedium xerophyticum possesses unilocular ovaries in contrast to the trilocular ones borne by the rest of the genus (Pfitzer, 1903; Atwood, 1984; Soto, Salazar, and Hagsater, 1990; Fig. 1). Paphiopedilum , the putative sister genus to Phragmipedium, always has unilocular ovaries in midsection, although basal and apical portions may show varying (and seemingly random) patterns of placental fusion (Atwood, 1984, p. 176). We disagree with Atwood's (1984, p. 186) opinion that because "intermediate conditions exist...emphasis on differences in locule number based on midsections is not justified." To the contrary, the cases of carpel fusion he documented do not represent intermediate character states because the taxonomically useful comparison is at midsection, where placentation (the character) is consistently either parietal or axile (two character states).
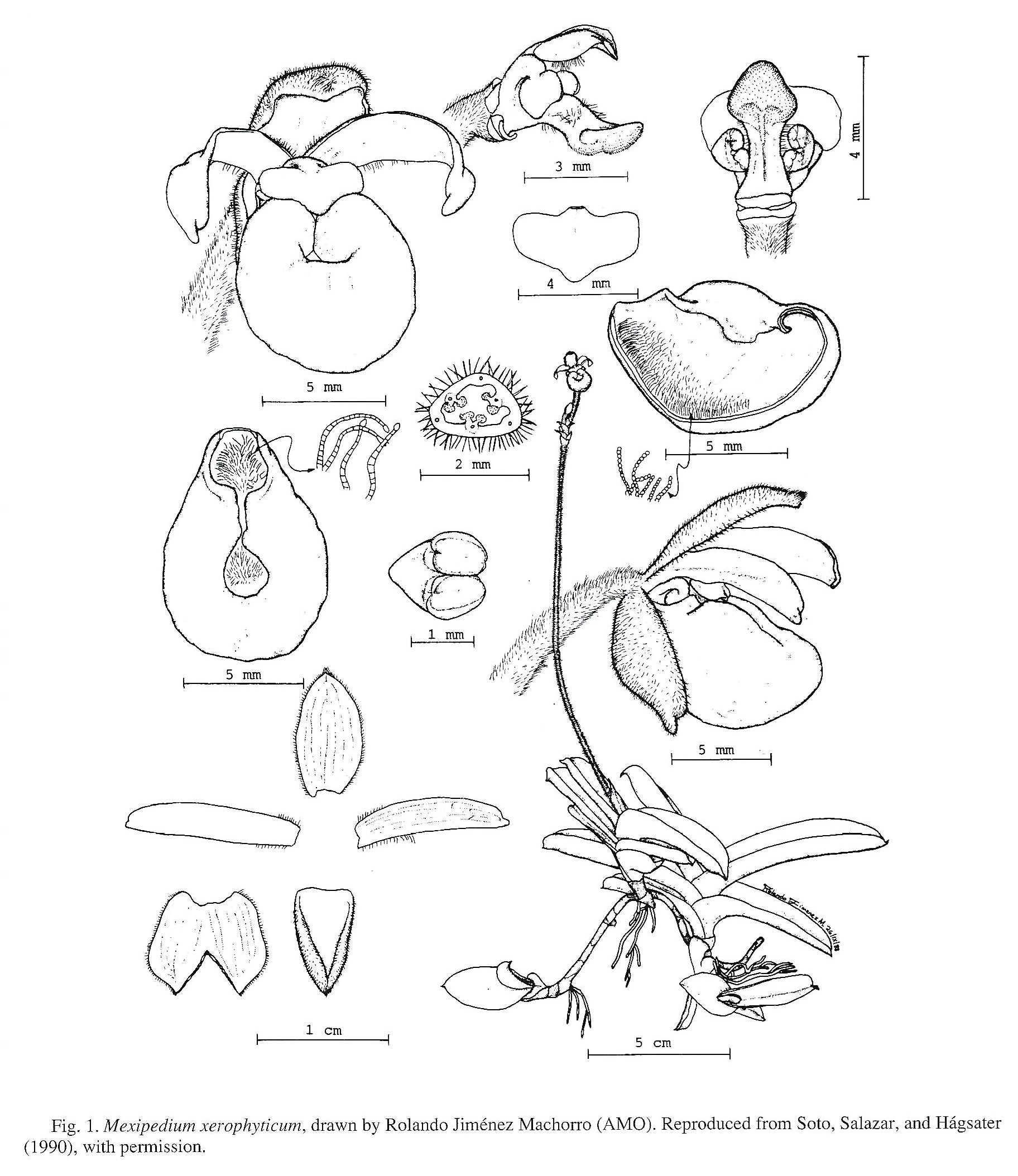
In summary, the generic assignment of Phragmipedium xerophyticum makes the taxonomic distinction between Paphiopedilum and Phragmipedium problematic. Rather than combining Phragmipedium with Paphiopedilum (which has nomenclatural priority), we propose a new, monotypic genus for Phragmipedium xerophyticum.
Mexipedium V. A. Albert & M. W. Chase, gen. nov. TYPE: Mexipedium xerophyticum (Soto , Salazar & Hagsater) V. A. Albert & M. W. Chase.
Genus Phragmipedium Rolfe similis, sed differt ovariis unilocularibus. Genus Paphiopedilum Pfitzer similis, sed differt racemis ramosis, sepalis cum aestivatione valvata.
Plants similar to genus Phragmipedium Rolfe and Paphiopedilum Pfitzer.
Inflorescences usually bearing two racemes, the apical one developing first, the basal one developing secondarily from the single, medial bract of the flowering scape;
raceme internodes abbreviated, bearing imbricating bracts.
Flowers borne successively.
Floral buds with valvate sepal aestivation.
Labellum calceolate and inflated.
Ovaries unilocular with parietal placentation.
Etymology : It is our pleasure to name this genus after its country of endemicity, Mexico.
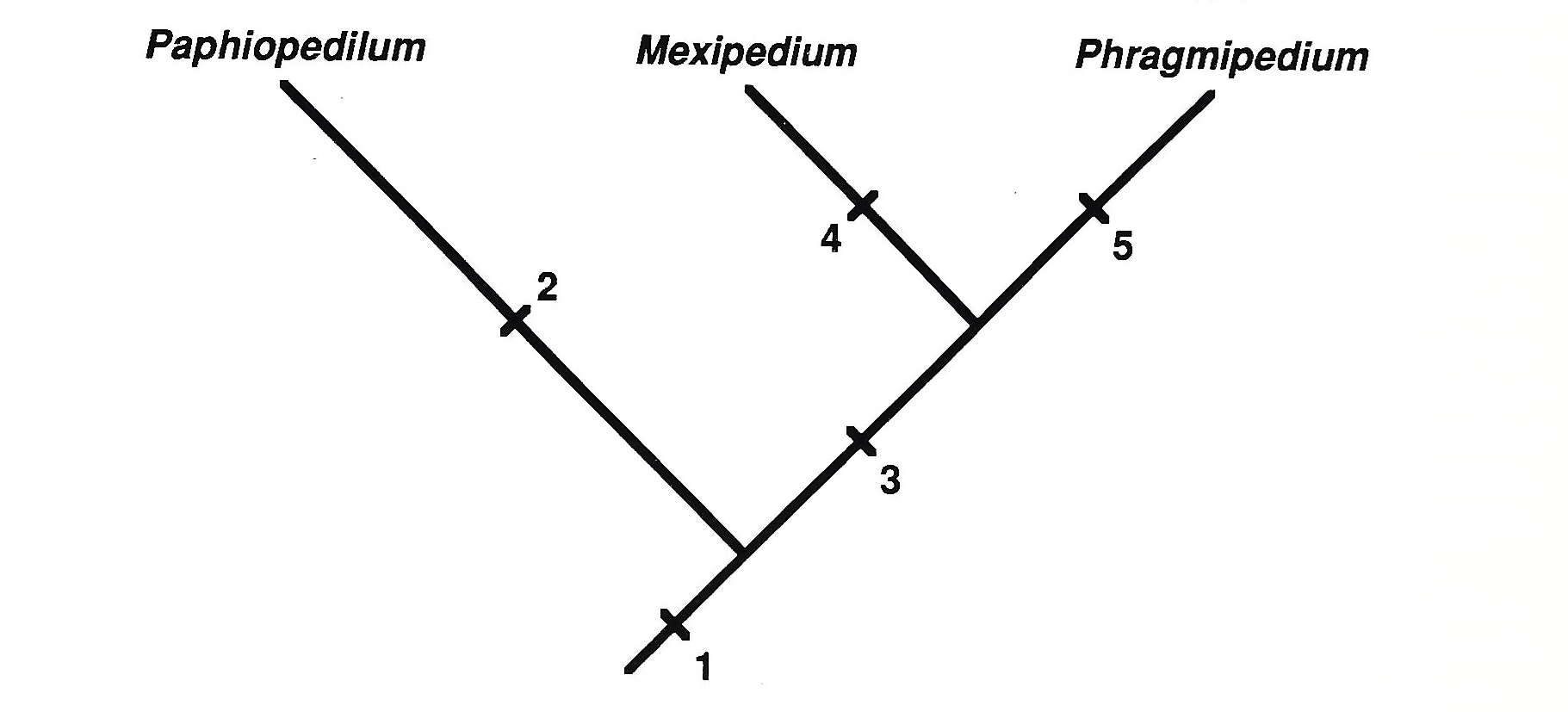
2. unbranched racemes, unilocular ovaries, imbricate sepal aestivation
3. [branched racemes] valvate sepal aestivation
4. unilocular ovaries
5. trilocular ovaries
Fig. 3. Phylogenetic key for the three genera of conduplicate-leaved slipper orchids. Diagnostic features for each lineage are indicated. For lineage (3), "branched racemes" is shown in brackets to reflect the polymorphism of this characteristic within Phragmipedium.
Mexipedium xerophyticum (Soto, Salazar, & Hagsater) V. A. Albert & M. W. Chase, comb. nov. Basionym: Phragmipedium xerophyticum Soto, Salazar, & Hagsater, Orquídea (Méx) 12:2. 1990. TYPE: MEXICO, Oaxaca: selvas de la vertiente del Golfo de México, 320 m s.n.m., vegetación xerofitica de Agave, Beaucarnea, Bursera simaruba, Plumeria y Pseudobombax ellipticum, en zona cárstica rodeada de selva alta perennifolia y encinares tropicales; hierba rupícola, escasa, flares blancas esfumadas de rosa, 6 septiembre 1988, G. A. Salazar 3740, M. A. Soto , E. Yáñez y H. Hernández (Holotype: AMO #11587; Isotype: K; photocopy of holotype seen).
Distribution: Endemic to Oaxaca, Mexico. Only seven clones are known, all from the type locality (intentionally unspecified for conservation purposes).
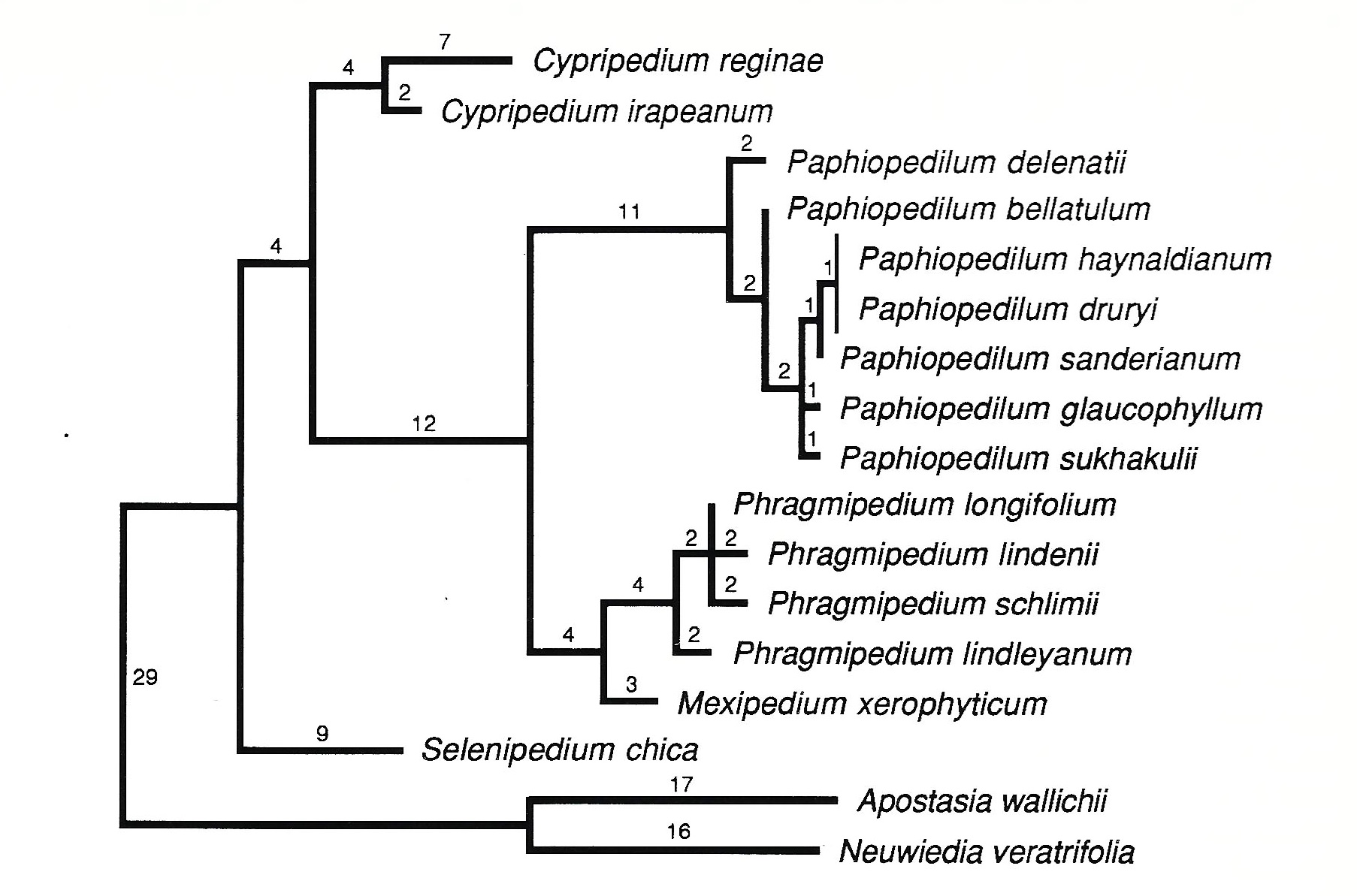
The recognition of Mexipedium stabilizes both Paphiopedilum and Phragmipedium by segregating a taxon with a problematic combination of character states (unilocular ovaries, branched racemes, and valvate sepal aestivation). This decision is supported by phylogenetic analysis of molecular, anatomical, and morphological data from representatives of the five genera of slipper orchids accepted here (V. A. Albert, unpubl.). Cypripedium, Mexipedium, Paphiopedilum, Phragmipedium, and Selenipedium are all monophyletic taxa (Fig. 2). The branch linking the conduplicate-leaved genera (Mexipedium, Paphiopedilum, and Phragmipedium) is well defined by twelve character-state changes. Similarly, the branch supporting Old World Paphiopedilum is marked by eleven changes. In contrast, the New World clade (Mexipedium plus Phragmipedium) is supported by only four changes, and Phragmipedium itself by four (Fig. 2). The composite branch length since common ancestry with Paphiopedilum is thus eight, which is comparable to the eleven changes supporting that genus. Additionally, Mexipedium has three unique changes (autapomorphies) assigned to its terminal branch, which is comparable to the four defining the entire Phragmipedium lineage (Fig. 2). These patterns of character support suggest that Mexipedium has undergone substantial morphological and molecular evolution in isolation from Phragmipedium, which is in keeping with its biogeographic distinction as the northernmost representative of the New World conduplicate-leaved slipper orchids.
A graphic "key" to Mexipedium, Paphiopedilum, and Phragmipedium is presented, with diagnostic features mapped onto the branching relationships hypothesized (Fig. 3).